Эксперименты с изменением параметров базовой модели видообразования.
Эта страница является продолжением и для ее понимания необходимо просмотреть часть 1,
описание
базовой модели эволюции
Частота мутаций.
Рассмотрим влияние частоты мутаций на ход эволюционного процесса. Напомним, что в базовой модели частота мутаций равнялась 0.001
Изменим это значения в 10 раз в сторону увеличения и уменьшения.
Частота мутаций (Pm) 0.01
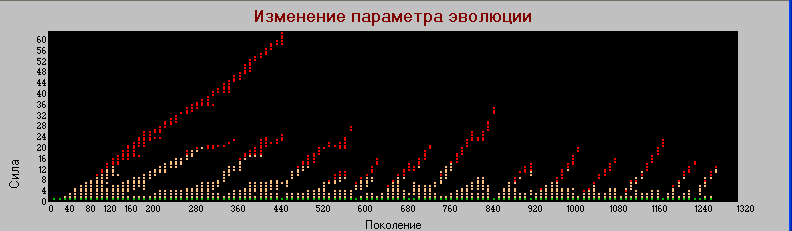
График «поколения» - «сила».
Эволюционный процесс происходит также как и и в базовой модели, но значительно быстрее и максимальное значение "силы" у первичных "гигантов" достигает значения 63 (в базовой модели - 39). Виды быстрее развиваются, но и быстрее вымирают. Так в базовой модели первичные "гиганты" вымирают после 1320 поколения, в то время как при Pm=0.01 где-то в районе 400-го поколения.
Но самое главное, что после 1380-го поколения вымирают все виды. Т.е. при высокой частоте мутаций система становится неустойчивой. Повидимому естественный отбор не успевает "обработать" быстро возникающие изменения.
Возникает вопрос, если процесс идет быстрее, не произойдет ли полное вымирание также и в базовой модели в последующих поколениях. Если умножить 1320 на 10, то где-то в районе поколения 14000 можно ждать катастрофы (напомним в базовой модели эксперимент был остановлен на поколении 8000). Был проведен эксперимент с базовой моделью до поколения 50000 и никаких признаков "катастрофы" не было обнаружено.
Частота мутаций (Pm) - 0.0001
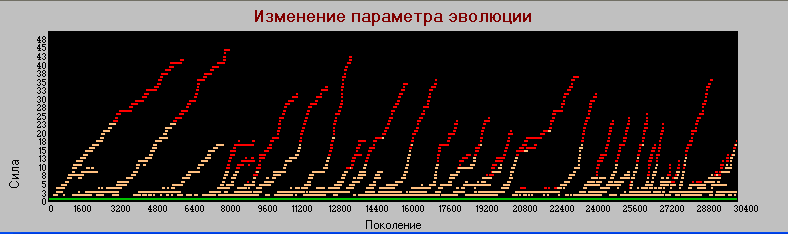
График 1 «поколения» - «сила». Pm=0.0001. Число поколений - 30 тыс.
Если предположить, что скорость эволюции пропорциональна частоте мутаций, то 30 тыс. поколений при РМ=0.0001 эквивалентно 3 тыс. при Pm=0.001
Ниже, для сравнения, приведены два графика
базовой модели (РМ=0.001)
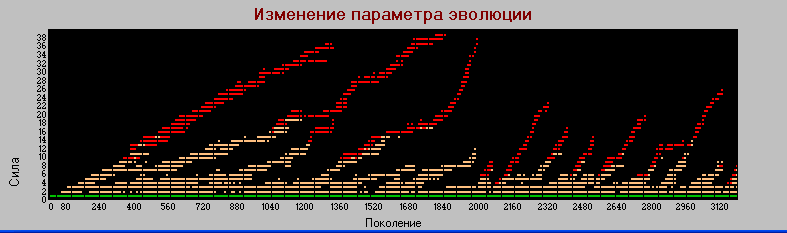
График 2 «поколения» - «сила» Pm=0.001 (базовый). Число поколений - 3 тыс.
Сравнивая графики 1 и 2 видим, что при уменьшении частоты мутаций, процесс эволюции замедляется (растягивается), однако пропорциональность замедления все же не прямая, а с неким коэффициентом меньшим единицы.
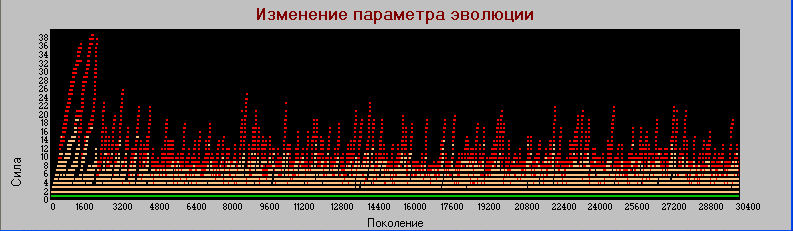
График 2а «поколения» - «сила» Pm=0.001 (базовый).Число поколений - 30 тыс.
График 2а эквивалентен по числу поколений графику 1. Видно, как увеличение частоты мутаций "сжимает" эволюционный процесс.
Из этого эксперимента можно сделать вывод, что изменение частоты мутаций влияет на скорость эволюционного процесса, но не на характер видообразования. Во всех случаях сохраняется циклический процесс видообразования.
В то же время высокая частота мутаций может привести к полному вымиранию всех видов.
"Калорийность" клетки.
Установим "калорийность" каждой клетки
k=4. Напомним, что в базовой модели эта величина была равна единице.
Частоту мутаций определим такую же как в базовой модели:
Pm=0.001.
В этом варианте предполагается более благоприятная среда для набора калорий особей.
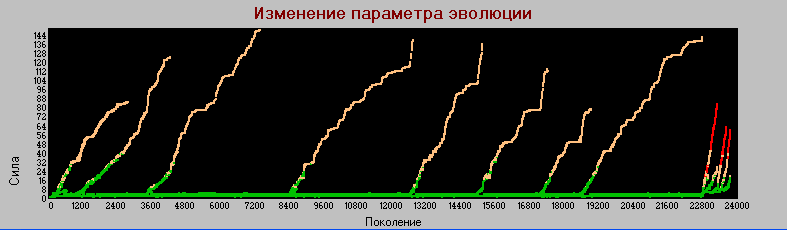
График 3а «поколения» - «сила» k=4.
Здесь видим, что видообразование происходит так же как и в предыдущих вариантах, т.е. циклическое появление и вымирание хищников при стабильном существовании основной линии травоядных, которые служат основой для пропитания хищников и материалом для их возникновения.
Однако существуют и отличия.
Стабильная линия травоядных имеет параметры сила, зона - (3,1) (в предыдущих экспериментах при k=1 эти значения были (1,1)). Что не удивительно при увеличении энергоемкости среды.
Циклы видообразования более четкие и редкие, так в базовой модели на интервале 5 тыс. поколений возникало более 27 циклов, причем некотрые из них "недоразвитые"и не совсем различимые, в то время как в данном варианте (k=4) на том же интервале таких циклов только три и они четко выражены. При этом максимальное значение силы равно 146 (в базовой - 39).
Однако в районе 22750-го поколения происходит качественное изменение процесса эволюции. У основной линии травоядных начинают быстро расти параметры "сила" и "зона", (в основном первый из них). При этом они остаются травоядными, благо энергоемкость среды это позволяет. В короткое время эти параметры достигают значений (10,2), система входит в неустойчивое состояние и в 23770-м поколении происходит полное вымирание.
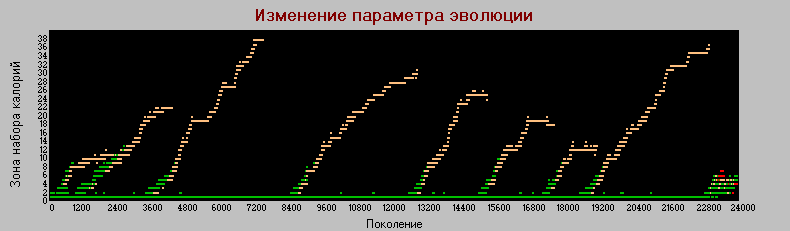
График 3б «поколения» - «зона» k=4.
Здесь также серьезное отличие от базового варианта, в котором различие между хищниками и травоядными по этому параметру наблюдались только на начальных циклах эволюции. До точки неустойчивости, разница очень четкая, наблюдается высокая корреляция с параметром силы. Т.е. в этом варианте четкий дизруптивный отбор идет сразу по двум признакам, в базовой модели это наблюдалось только по признаку "сила".
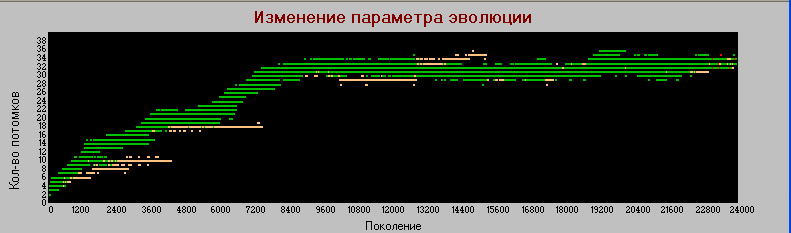
График 3в «поколения» - «плодовитость» k=4.
По этому параметру тоже наблюдаются некоторые различия. В базовом варианте «плодовитость» хотя и с замедлением, но растет постоянно на всем протяжении эксперимента (50 тыс. поколений). В настоящем варианте рост практически прекращается где-то после 7 тыс. поколений
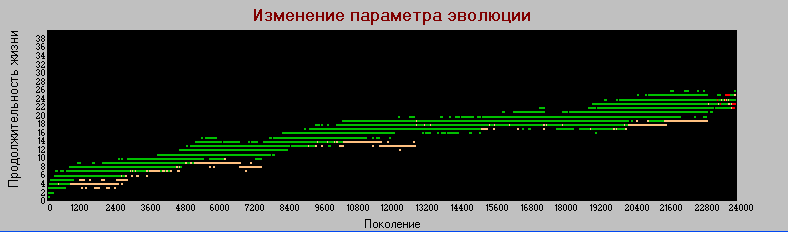
График 3г «поколения» - «долголетие» k=4.
А вот здесь наоборот, рост параметра хотя и медленно, но увеличивается. В базовом варианте рост долголетия прекращается после поколения 1500.
Есть еще одно существенное отличие данного варианта от базового. Это количественное соотношение между хищниками и травоядными. В данном варианте (до неустойчивого участка) процент хищников составляет 3 - 4%, в то время как в базовой модели - примерно 25%. А на неустойчивом участке данного варианта это соотношение доходит до 50%
Увеличим значение калорийности среды еще в два раза k=8
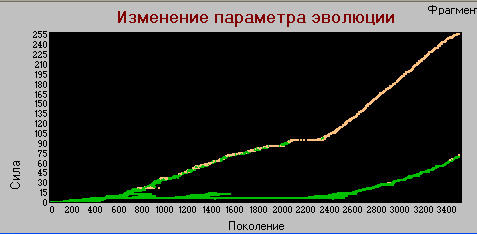
График 3а «поколения» - «сила» k=4.
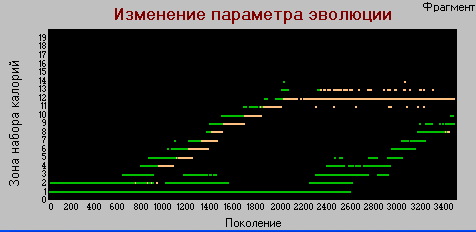
График 3б «поколения» - «зона» k=4.
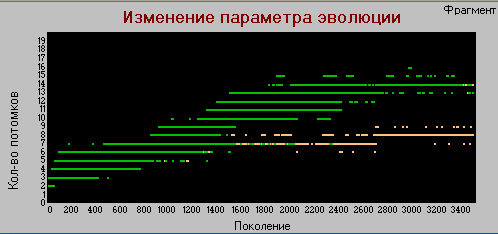 График 3в «поколения» - «плодовитость» k=4.
График 3в «поколения» - «плодовитость» k=4.
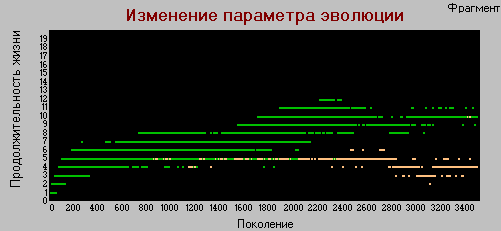 График 3г «поколения» - «долголетие» k=4.
График 3г «поколения» - «долголетие» k=4.
В этом варианте формы графиков эволюции практически повторяют аналогичные формы предыдущего варианта
k=4 до 1200-го поколения. Только здесь они растянуты до поколения 3500. При этом развитие количественное значений признака "сила" достигает значительно большего значения - 255 (вариант k=4 - 146) и за более короткое время.
Однако на 3500-м поколении произошло полное вымирание всех видов, т.е. значительно быстрее чем в предыдущем варианте (почти в 7 раз)
Так что же получается, чем больше энергоемкость среды, тем неусточевее биоценоз ? Разве это не противоречит всему, что мы видим в жизни ?
Ну, до жизни этой модели далеко, а видим мы только то, что бурное развитие, за которым "не успевает" естественный отбор, действительно может приводить к катастрофическим последствиям. И действительно большая энергоемкость среды такому бурному развитию способствует.



