Взгляни на первую лужу – и в ней найдешь гада,
который иеройством своим всех прочих гадов превосходит и затемняет.
М.Е. Салтыков-Щедрин
История одного города.
Часть 1. Базовая модель видообразования.
Согласно общепринятой точке зрения, в основе видообразования лежит принцип
дивергенции. Именно, в результате дивергенции увеличивается число видов. Термин
«дивергенция признаков» введён Ч. Дарвином (1859).
Видообразование в однородной среде, т.е.видообразование без физических барьеров, называют
>симпатрическим видообразованием.
В отличии от
аллопатрического видообразования , где исходный вид разделяется на части физическими барьерами — проливом, горным хребтом и т. п. и в изолированных популяциях постепенно накапливаются генетические различия, которые со временем становятся столь значительными, что делают невозможным скрещивание представителей двух разобщенных популяций, даже если разделявший их барьер исчезнет и они снова встретятся.
И поскольку они не могут скрещиваться - два вида существуют вместе.
А почему они будут сосуществовать вместе? Ведь у Дарвина написано что самая свирепая конкуренция как раз между близкими видами. Или они так далеко разошлись, что уже и не конкурируют?
И вообще, рушатся горы, долины, испаряются проливы - жуткая картина.
Ну и наконец, на раннем этапе развития жизни, когда не было полового размножения и соответственно изолирующего скрещивания?
Или тогда не было видообразования?
Приведем несколько определений:
Дивергенция (от средневекового лат. divergo — отклоняюсь), в биологии — расхождение признаков и свойств у первоначально близких групп организмов в ходе эволюции. Результат обитания в разных условиях и неодинаково направленного естественного отбора.
Универсальная энциклопедия.
Если вид занимает обширный ареал и приспосабливается к разным экологическим условиям, то возникает Дивергенция, выражающаяся в появлении каких-либо различий между первоначально сходными популяциями и обусловленная неизбежно несколько неодинаковым направлением естественного отбора в разных частях ареала вида.
А. В. Яблоков. (БСЭ)
Дивергенция (расхождение признаков) двух или нескольких таксонов, возникающих от общего предка, обусловлена различиями в условиях среды………
А. С. Северцов.
Как видим, во всех этих определениях присутствует положение о предварительной изоляции частей популяции и их более или менее автономного развития либо в разных условиях среды, либо в разных частях ареала.
Единственное четкое указание, даже не о возможности, а о неизбежности дивергенции, без всякой изоляции, я встретил у М.Е. Салтыкова-Щедрина (см. эпиграф).
Действительно, возникновение «иеройского гада» может произойти только в среде «гадов неиеройских», иначе кто же будет им восторгаться и трепетать и главное, предоставлять ему пропитание собственной шкурой ?
Для проверки этого положения была разработана
компьютерная модель эволюции, реализующая так называемое имитационное моделирование, его иногда называют
машинным моделированием.
Позволим себе еще одно определение:
Имитационное моделирование — это метод, позволяющий строить модели, описывающие процессы так, как они проходили бы в действительности. Такую модель можно «проиграть» во времени как для одного испытания, так и заданного их множества. При этом результаты будут определяться случайным характером процессов. По этим данным можно получить достаточно устойчивую статистику.
Цель данной работы - не доказать то-то и то-то, а посмотреть что будет.
Теперь к делу.
Описание модели эволюции.
Модель имитирует эволюционный процесс на популяциях «организмов» без полового размножения. Это, кроме упрощения модели, позволяет имитировать механизм
естественного отбора в чистом виде. Предполагается что естественный отбор имел место и до возникновения механизма скрещивания.
Процесс
биологической эволюции моделируется на квадратной сетке размером
n x
n. Каждая клетка сетки имеет условную «калорийность» (k) . На сетке могут быть расположены «особи», таким образом, что в каждой клетке может быть расположена только одна особь.
Каждая особь имеет следующие числовые признаки:
- «сила» (S), особь с большим значением силы может «поглотить» любую особь с меньшим ее значением,
- «зона набора калорий» (L) или кратко «зона», определяющий величину пространства на котором особь может поглощать пустые клетки и других особей (не смог подобрать лучшего названия, можно назвать «зона охоты»),
- «плодовитость» (C), число потомков особи,
- «долголетие» (V), определяющее число циклов охота-размножение особи.
«Поколением» считается перебор всех особей, размешенных на сетке, с проведением для каждой из них цикла охота-размножение.
Опишем стандартный цикл каждой (текущей) особи.
"Охота" (набор калорий) – производится случайный выбор клетки в квадрате со стороной двойной «зоны набора калорий» (2*L ). Если клетка пуста – в актив особи записывается калорийность пустой клетки k. Если клетка занята другой особью – сравниваются их силы и если сила потенциальной «жертвы» меньше, то она поглощается и в актив особи записывается калорийность жертвы равная сумме «силы» и «зоны» жертвы плюс калорийность клетки, в которой эта жертва находится - (S+L+k). После чего жертва уничтожается.
Если клетка занята особью с признаком силы не меньшей чем у текущей, то в актив текущей особи ничего не записывается .
Подобным образом случайно выбираются остальные L-1 клеток (по величине «зоны набора калорий» текущей особи). К сумме приобретенных калорий прибавляются также калории клетки, в которой расположена текущая особь.
После каждого шага охоты подсчитывается энергетический баланс. Если сумма приобретенных калорий не меньше суммы признаков текущей особи S+L, то охота прекращается и особь приступает к размножению, в противном случае она уничтожается (умирает от истощения).
Заметим, что здесь сумма признаков S+L как бы символизирует физические размеры особи.
Размножение – производится случайный выбор клетки в квадрате с двойной стороной значения признака «плодовитость» (C). Если клетка свободна, в ней размещается потомок, в противном случае размещение не происходит. Подобным образом производится размещение (или неразмещение) всех C потомков.
Мутации – при размещении потомка в клетке производится мутация для каждого признака с вероятностью Pm, которая заключается в изменении значения признака на единицу в ту или другую сторону, с равной вероятностью. Если при этом признак принимает нулевое значение, потомок уничтожается как нежизнеспособный.
По окончанию обработки поколения (т.е. всех расположенных на сетке особей, кроме новорожденных), высвечиваются данные определяющие состояние эволюционной системы – расположение особей на сетке, таблица видов и векторное поле, показывающее положение каждого вида в пространстве двух выбранных признаков.
Уточним понятие вида в данной модели. В биологии это группа особей с общими морфо-физиологическими, способная к взаимному скрещиванию, дающему в ряду поколений плодовитое потомство.
В нашей модели скрещивание отсутствует, поэтому для простоты будем пользоваться следующим определением: под видом понимается группа особей, отличающаяся от других, любым значением хотя бы одного признака.
Предполагается, что в результате эволюции, количественные значения признаков видов будут расходиться, с потерей промежуточных значений (
дизруптивный отбор ).
В результате экспериментов с моделью хотелось узнать действительно ли будет иметь место
дизруптивный отбор и если да, то по каким признакам и насколько устойчивыми являются конфигурации, получаемые в процессе эволюции.
Результаты каждого десятого поколения эволюционного моделирования (таблица видов) записываются в базу данных, что позволяет проводить их анализ по окончании эксперимента.
Базовый эксперимент.
Размер сетки равен n=100, т.е. моделирование производится на сетке 100х100 клеток.
"Калорийность" каждой клетки k=1
В середине сетки размещается исходная особь, имеющая следующие значения признаков:
- «сила» S= 1,
- «зона набора калорий» L=1,
- «плодовитость» C=2,
- «долголетие» D=1
Будем называть последовательность значений признаков (S,L,C,D) векторм признаков вида.
Таким образом исходная особь имеет вектор (1,1,2,1).
Вероятность мутации каждого признака в эксперименте Pm=0.001.
Эксперимент был остановлен по достижении 8000-го поколения.
Рассмотрим некоторые промежуточные результаты:
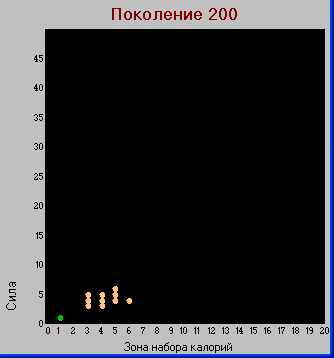
Поколение 200.
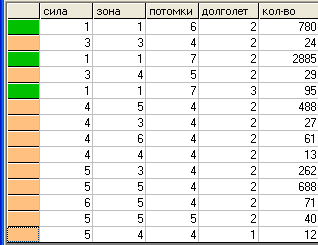
Таблица "видов".
В
таблице видов показаны все варианты, возникшие в процессе эволюции к данному поколению. Здесь видно, что различия имеют место только по первым трем признакам. По признаку «долголетие» расхождений практически нет, да и изменился он не сильно.
Полученные виды различаются по калорийному балансу, так для калорийности каждой клетки поля k=1 только виды (1,1,х,х) могут существовать потребляя только пустые клетки (им нужно 1+1 = 2 калории, что выполняется при L=1 плюс клетка, в которой особь находится).
Будем называть виды первого типа "травоядными" (если точно – клеткоядными), а второго "хищниками" (хотя на самом деле они всеядные), соответственно первые имеют зеленый цвет, вторые желтый.
Если сумма калорий, необходимых для выживания особи равна S+L, то травоядные определяются условием: S+L <= k*(L +1). Поэтому для калорийности клетки k=1, единственный вариант для травоядных: S=1; L=1. Во всех остальных вариантах особь становится или нежизнеспособной или резко уменьшается вероятность ее выживания или она переходит в группу хищников
Т.е. в результате эволюционного процесса, получено не только разнообразие видов, но и некое подобие трофической цепи.
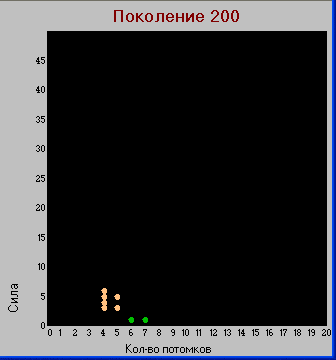
Для более наглядного отображения ожидаемого расхождения признаков будем изображать виды в двумерных подпространствах наиболее изменяемы признаков:
«сила» (осьY) - «зона» (осьX) и
«сила» (осьY) - «плодовитость» (осьX).
При таком представлении, каждый вид изображается точкой с координатами X,Y
Здесь видно, что по признакам «сила» и «зона» происходит разделение – у хищников эти признаки больше чем у травоядных, а вот по количеству потомков – наоборот.
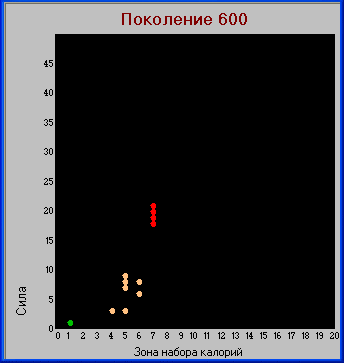
Поколение 600.
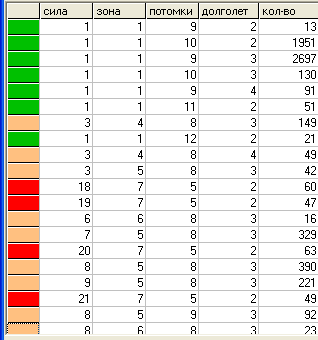 Таблица "видов".
Таблица "видов".
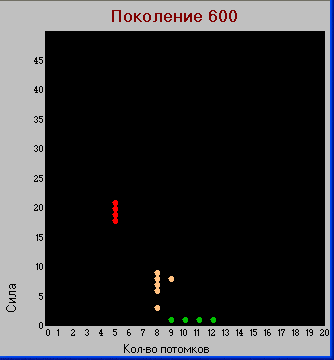
Разнообразие видов значительно увеличилось. Появились виды, для выживания которых не хватает не только количества доступных им пустых клеток, но и находящихся в них травоядных. Т.е. для выживания они должны поглощать не только пустые клетки и травоядных, но и некоторое количество хищников с меньшей «силой». Будем называть такие виды хищниками второго уровня и присвоим им красный цвет.
Хищники второго уровня определяются условием: S+L > k*(L +1)+2*L, здесь коэфф. 2 определяет калорийность травоядной особи в данной модели («сила»=1+ «зона»=1).
В остальных случаях имеем хищника первого уровня.
В этом поколении уже появляется четкое разделение на описанные выше группы видов, с отсутствием промежуточных вариантов, т.е. налицо
дизруптивный отбор. Также прослеживается обратная зависимость между параметрами «сила» и «плодовитость» (заметим, что подобная зависимость имеется и в живой природе - у более сильных видов, как правило, меньше потомков).
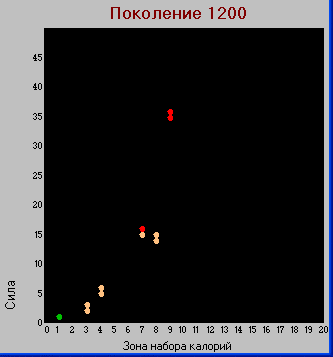
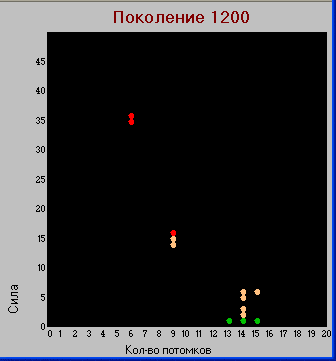
Поколение 1200.
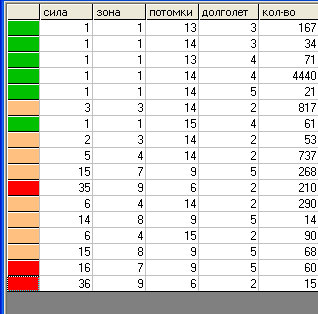 Таблица "видов".
Таблица "видов". Здесь заметно дальнейшее развитие хищников обеих групп, в основном, по параметру «сила». По количеству потомков эти группы не развиваются. Травоядные продолжают увеличивать значение параметра «плодовитость».
В тоже время видим, что в результате эволюции из травоядных появляется новая группа хищников, в отличие от предыдущих обладающая большим количеством потомков вытеснившая первоначальных "гигантов".
Диаграммы дивергентной эволюции видов
Для анализа динамики эволюции на всем протяжении эксперимента воспользуемся графиками «Изменение параметра эволюции». В каждом из этих графиков, по оси X расположены поколения (с интервалом 10) а по оси Y значения одного из параметров для всех видов данного поколения. Т.е. каждому поколению (ось X) соответствует m точек оси Y, где m – количество видов в данном поколении.
Рассмотрим график
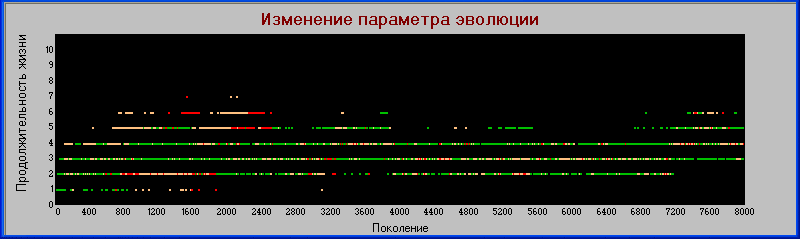
«поколения» - «сила».
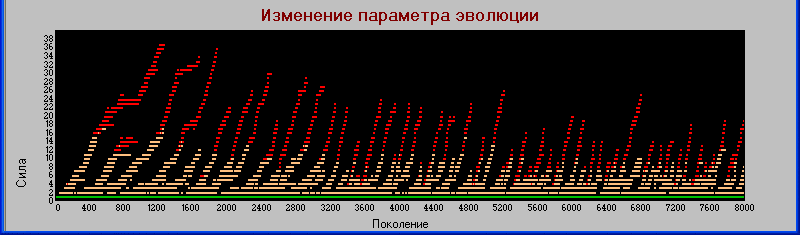
Здесь видно, что эволюция по этому параметру происходит циклически, с созданием трофических цепей, вымиранием хищников второго (и иногда даже первого) уровня и последующим эволюционированием хищников первого уровня в сторону силы до появления новых «гигантов».
В то же время травоядные (на графике они изображены в виде зеленой полосы около оси Х) дают материал для появления и развития новых хищников первого уровня, которые после вымирания «гигантов» заполняют освобождающуюся трофическую нишу и сами начинают развиваться в сторону гигантизма.
В данной модели, травоядные развиваются преимущественно по параметру «плодовитость», тогда как хищники, в основном, по параметру «сила» и в меньшей степени по признаку «зона».
Однако, на каждом последующем цикле эволюции вновь появляющиеся хищники получают от травоядных по наследству увеличенное значение параметра «плодовитость». Поэтому, если на первом цикле эволюции хищники и травоядные расходились по обоим признакам «сила» и «плодовитость», то в последующих циклах разделение по потомкам резко уменьшилось, хотя и не исчезло совсем.
Однако увеличение плодовитости хищников не остается без последствий - они "мельчают".
И самое главное - это увеличение плодовитости происходит не потому что это увеличивает их жизнеспособность (из диаграммы видно что совсем наоборот - первичные гиганты существовали значительно дольше), а просто они изначально были созданы такими.
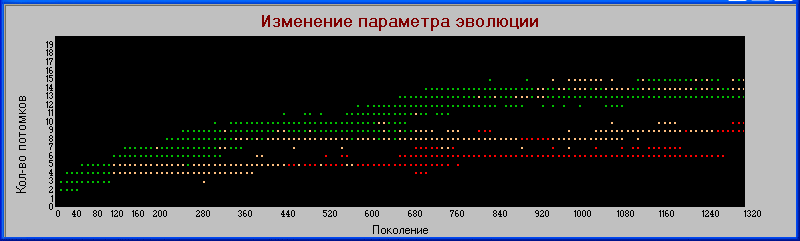
Иллюстрацией этому служит график
«поколения» - «плодовитость».
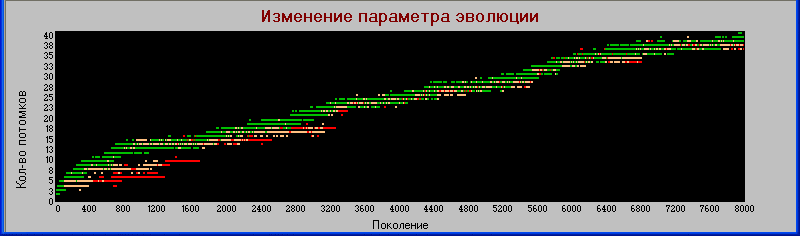
В начале эволюции довольно четкое разделение: хищники имеют меньшую плодовитость. Но потом наступает перемешивание, хотя некоторое разделение все же можно заметить.
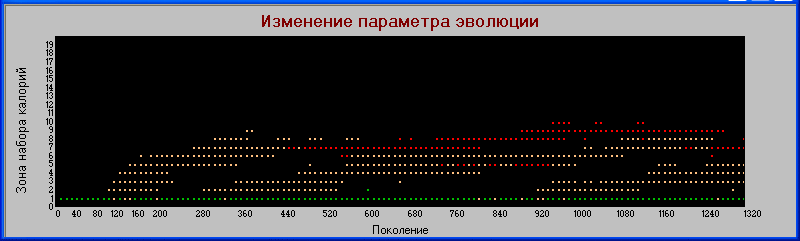
График
«поколения» - «зона».
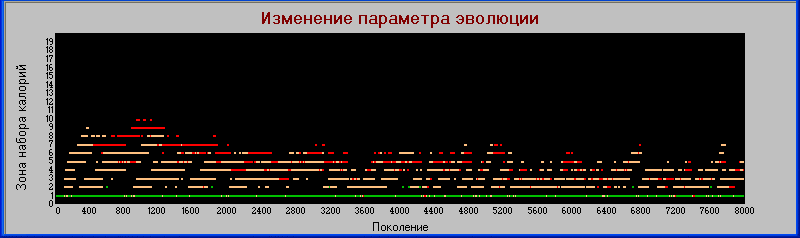
По признаку «зона» заметно расхождение между хищниками первого и второго уровней, увеличение «силы» совпадает с увеличением зоны, но только в начальных циклах. В дальнейшем это расхождение нивелируется.
На начальном этапе эволюции видна тенденция к увеличению значения признака "зоны", однако далее это значение не только не увеличивается, но даже падает. По видимому это связано с параллельным увеличением плодовитости, которое становится основным фактором выживания вида.
График
«поколения» - «долголетие».
Признак «долголетие» в процессе эволюции как-то незаметен. На начальном этапе его значение довольно быстро возрастает и далее постоянно находится в интервале 2-5. Остановка роста этого параметра связана с развитием более эффективного признака - "плодовитости".
Можно предположить, что вымирание «гигантов» обусловлено развитием хищников первого уровня, которые хотя и являются их пищей, но в свою очередь уничтожая большое количество травоядых, являются более живучими, в первую очередь за счет большего количества потомков.
С другой стороны в последующих циклах количество потомков у всех видов выравнивается, однако вымирание «гигантов» продолжается.
Можно рассмотреть и обратное предположение: интенсивное развитие признака «сила» требует для выживания большего количества калорий, вероятность получения которых падает. Это, в свою очередь, позволяет развиваться хищникам первого уровня, что еще более ухудшает ситуацию для «гигантов».
Но наверное оба эти фактора действуют одновременно.
В общем виде механизм возникновения трофических цепочек на однородной среде можно представить следующим образом. Сначала происходит развитие «травоядных» в сторону увеличения потомков. По мере достижения ими определенной плотности создается среда, в которой делается возможным развитие особей для выживания которых недостаточно пустых клеток, т.е. происходит возникновение «хищников».
После этого, первые начинают интенсивно развиваться в сторону увеличения потомства, вторые в сторону «силы» и «зоны». Особенно четко это видно на первом цикле эволюции.
Таким образом, видим что даже на столь примитивной модели имеет место
дивергентная эволюция и дизруптивный отбор.
Замечу, что при создании модели все четыре признака выбирались «от балды» и оказалось что признак «долголетие» в дивергентной эволюции никак себя не проявляет, являясь придатком признака «плодовитость». Наверно для него модель слишком примитивна.
Неожиданным, хотя и желанным был эффект образования и вымирания гигантов (конечно в голове вертелись динозавры, что за эволюция без них).
Проблема устойчивости видов.
И все-таки, почему гиганты бурно развиваются и затем внезапно вымирают ? Является ли устойчивой «экосистема» до вымирания гигантов или наоборот – вид давно прошел точку неустойчивости и только развитие какое-то время поддерживает его существование?
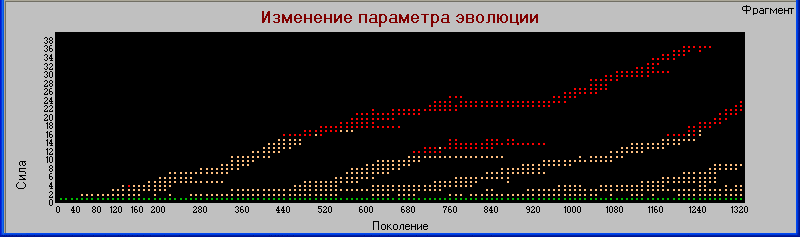
Прервем эксперимент на 2902 поколении, это как раз место, близкое к вымиранию гигантов в одном из циклов. Перекрываем мутацию и запускаем модель дальше до поколения 8000.
Результат выводим на график «поколения» - «сила».
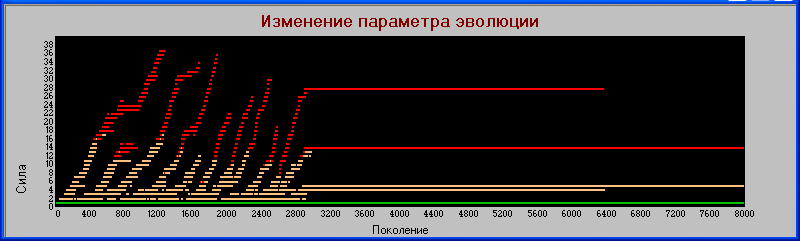
Здесь видно, что система продолжает быть устойчивой достаточно продолжительное время. И хотя гиганты все же вымирают, просуществовав более 3000 поколений, (напомним, что в процессе эволюции им осталось жизни всего несколько поколений) тем не менее столь долгое время просуществовала достаточно сложная конструкция из пяти видов с тремя уровнями трофической цепи.
Т.е. причиной вымирания является именно процесс эволюции, который здесь выступает в роли Тараса Бульбы ("Я тебя породил, я тебя и убью").
Заметим, что вымирание гигантов произошло одновременно с гибелью одного из видов хищников первого уровня.
Получается, что механизм эволюции (изменчивость плюс естественный отбор) вполне может создавать сложные, устойчивые конструкции и в то же время приводить к внезапным катастрофам
Иными словами, для объяснения вымирания видов необязательно искать внешние причины типа метеоритов,
вымирание может являться следствием самой эволюции.
Лирическое отступление.
Так отчего же вымерли динозавры?
Сейчас, по крайней мере в популярных источниках, практически доказанной причиной их гибели считается падение метеорита. То есть, если бы он не упал, то динозавры по сю пору гуляли бы, по крайней мере, в Африке. А вот их современников крокодилов метеорит не убил, не заметил что ли?
Когда-то, гипотез вымирания динозавров было много и изменение погоды (а крокодилы, крокодилы !!!) и то что мелкие, но вредные млекопитающие ели динозаврьи яйца и трава ядовитая появилась и пр. Жаль что в ту пору не было человека - точно бы свалили на него.
Среди них как-то незаметно проскочила мысль о том, что вымирание динозавров и не только их, является следствием самого процесса эволюции и без всяких внешних причин.
Однако эта гипотеза, не получила особого распространения. Причиной тому, повидимому распространенное мнение оголтелых дарвинистов, которые считают что естественный отбор все устраивает наилучшим образом и не может приводить к массовым вымираниям целых видов и даже таксонов. Кстати, массовые вымирания были и до динозавров, это сколько же надо метеоритов. Хотя, конечно, у Бога недостатка в инвентаре не бывает.
Это напоминает недавние заклинания наших экономических либералов, что «невидимая рука рынка» все всегда устраивает наилучшим образом. Хотя, как известно, кризисы были, есть и будут.
Описанный выше эксперимент конечно не является доказательством чего-нибудь, но возможно способствует появлению мыслей отличных от так называемого мэйнстрима.
А динозавры, Бог знает от чего вымерли.
На этой мажорной ноте пока и закончим.
Часть 2 Эксперименты с изменением параметров базовой модели видообразования.
В части 2 описаны эксперименты с изменением некоторых параметров модели, в частности частоты мутаций и "калорийности" среды (клеток).
Дальнейшие эксперименты с данной моделью будут изложены на этом сайте.
Работы на тему видообразования:
Дупликационная модель биологической эволюции